|
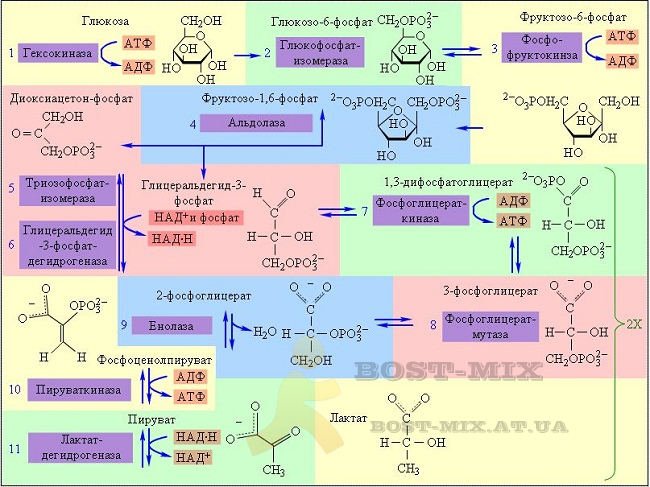
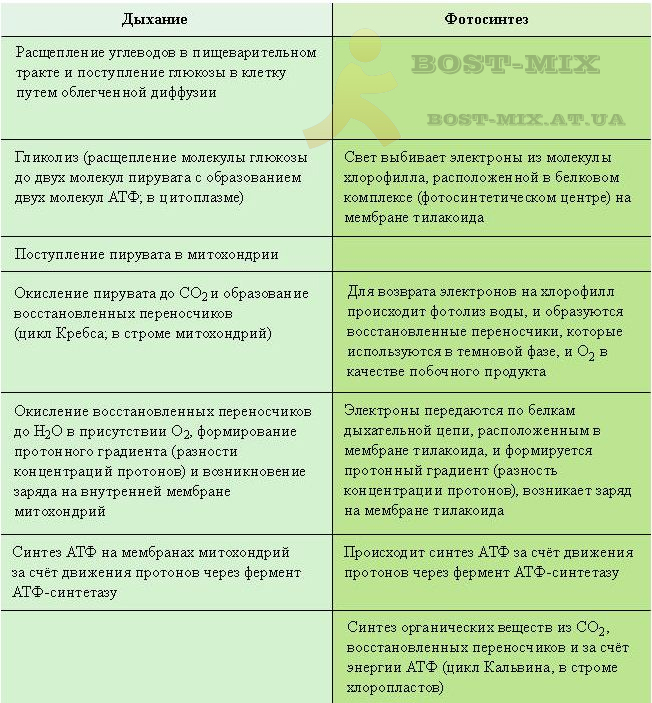
В отсутствие кислорода глюкоза в клетке окисляется лишь частично, не до углекислого газа, а лишь до пировиноградкой кислоты (пирувата). Этот процесс называют гликолизом. (У организмов, лишённых возможности в дальнейшем доокислять пируват (анаэробные бактерии), он может превращаться в другие не полностью окисленные продукты: молочную кислоту, этиловый спирт, уксусную кислоту, ацетон и др.: эти процессы называют брожением). В ходе гликолиза образуется (в пересчете на молекулу глюкозы) 2 молекулы АТФ и две молекулы восстановленного переносчика (НАД∙Н – никотинамидадениндинуклеотид восстановленный).
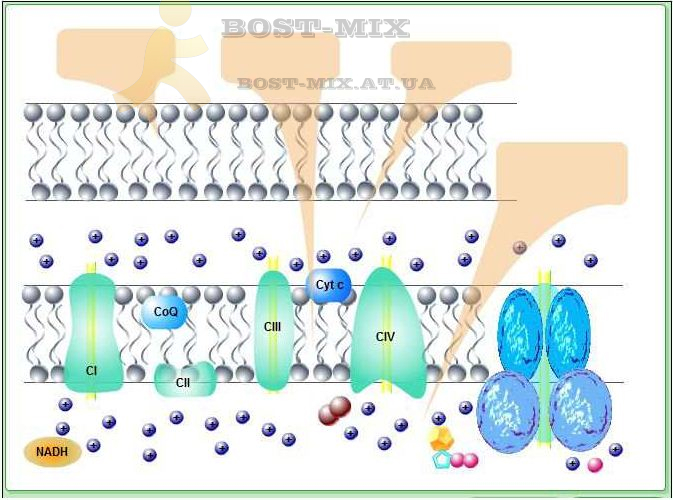
В присутствии кислорода процесс окисления глюкозы идет до конца (аэробное дыхание). В нем выделяют три основные стадии: окисление пирувата, цикл Кребса и окислительное фосфорилирование. Первые два процесса протекают у эукариот в матриксе митохондрий. Пируват, окисляясь и теряя молекулу углекислого газа, переносится в виде ацетил-кофермента А в цикл трикарбоновых кислот (цикл Кребса). В ходе одного оборота цикла все атомы углерода, входившие в молекулу пирувата, доокисляются до СО2 с образованием (в пересчете на молекулу глюкозы) двух молекулы ГТФ (что энергетически тождественно двум молекулам АТФ), и 8 молекул восстановленных переносчиков – 6 НАД∙Н и 2 ФАД∙Н2. Основное количество молекул АТФ (максимально – до 34 в пересчете на молекулу глюкозы) вырабатывается по способу окислительного фосфорилирования на последней стадии клеточного дыхания: в электронтранспортной цепи. Здесь происходит окисление НАД∙Н и ФАД∙Н2, восстановленных в процессах гликолиза, β-окисления жирных кислот, цикла Кребса и других процессов. Энергия, выделяющаяся в ходе этих реакций, благодаря цепи переносчиков электронов, локализованной во внутренней мембране митохондрий (у прокариот — в цитоплазматической мембране), преобразуется в трансмембранный протонный потенциал (разность зарядов на внутренней мембране митохондрий). Фермент АТФ-синтаза использует этот потенциал для синтеза АТФ, преобразуя его энергию в энергию химических связей. Подсчитано, что молекула НАД∙Н может дать в ходе этого процесса три молекулы АТФ, ФАД∙Н2 – две. Таким образом, аэробное дыхание энергетически гораздо выгоднее
гликолиза.
Конечным акцептором электрона в дыхательной цепи большинства организмов является кислород (его недостаток, например, в скелетных мышцах во время активной работы, ингибирует все фазы аэробного дыхания в клетке, вынуждая ее ограничиться гликолизом, сопровождающимся накоплением образованной из пирувата молочной кислоты). У некоторых хемосинтетических бактерий в электронтранспортной цепи вместо кислорода используется другой конечный акцептор (трёхвалентное железо, нитрат- или сульфат-анион). Хемосинтез осуществляется только бактериями, которые таким образом играют важную роль в биогеохимических циклах серы, азота и железа. Денитрификация – один из типов анаэробного дыхания – является одним из источников парниковых газов, железобактерии принимают участие в образовании железомарганцевых конкреций. Бактерии-хемоавтотрофы способны усваивать CO2 как единственный источник углерода за счёт энергии окисления неорганических соединений.
|