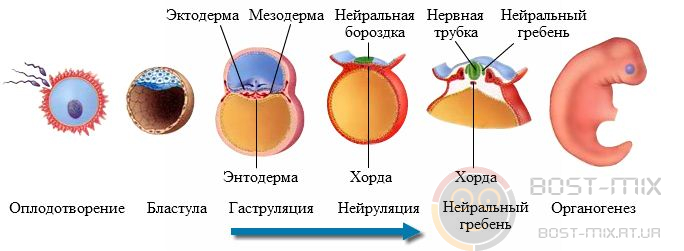
Онтогенезом называется индивидуальное развитие организма от оплодотворения до смерти. Термин был впервые введен У многоклеточных животных онтогенез делится на два периода:
В эмбриональном периоде выделяют три основных этапа: дробление, гаструляцию и первичный органогенез. Дробление – ряд последовательных митотических делений оплодотворённого или инициированного к развитию яйца. Дробление представляет собой первый период эмбрионального развития, который присутствует в онтогенезе всех многоклеточных животных и приводит к образованию зародыша: бластулы (однослойный полый шар из клеток) или, гораздо реже – морулы (шарообразная масса клеток без полости внутри). При этом масса зародыша и его объем не меняются, то есть они остаются такими же, как у зиготы, а яйцо разделяется на все более мелкие клетки – бластомеры. После каждого деления дробления Тип дробления зависит от количества желтка и его расположения в яйце. Если желтка мало и он равномерно распределён в цитоплазме (изолецитальные яйца: иглокожие, плоские черви, млекопитающие), то дробление протекает по типу полного равномерного: бластомеры одинаковы по размерам, дробится все яйцо. Если желток распределён неравномерно (телолецитальные яйца: амфибии), то дробление протекает по типу полного неравномерного: бластомеры – разной величины, те, которые содержат желток – крупнее, яйцо дробится целиком. При неполном дроблении желтка в яйцах настолько много, что борозды дробления не могут разделить его целиком. Дробление яйца, у которого дробится только сконцентрированная на анимальном полюсе «шапочка» цитоплазмы, где находится ядро зиготы, называется неполным дискоидальным (телолецитальные яйца: пресмыкающиеся, птицы). При неполном поверхностном дроблении в глубине желтка происходят первые синхронные ядерные деления, не сопровождающиеся образованием межклеточных границ. Ядра, окружённые небольшим количеством цитоплазмы, равномерно распределяются в желтке. Когда их становится достаточно много, они мигрируют в цитоплазму, где затем после образования межклеточных границ возникает бластодерма (центролецитальные яйца: насекомые). Гаструляция – процесс разделения зародыша на зародышевые листки. В ходе гаструляции клетки зародыша практически не растут, происходит активное передвижение клеточных масс (морфогенетические движения); двухслойный зародыш (гаструла) образуется путем впячивания участка бластулы, обрастания одного слоя другим или путем выселения клеток. Из наружного слоя клеток, эктодермы впоследствии закладывается эпителий кожных покровов, ротовой и анальной полостей, а также нервная трубка и зачатки органов чувств. Из внутреннего слоя, энтодермы, образуется эпителиальная выстилка пищеварительного тракта, органов дыхания, а также железистый эпителий пищеварительных желез (поджелудочной, слюнных, печени). Первичный органогенез – процесс образования комплекса осевых органов. В разных группах животных этот процесс характеризуется своими особенностями. Например, у хордовых на этом этапе происходит закладка нервной трубки (процесс нейруляции, стадия носит название нейрулы), хорды и кишечной трубки. При этом образуется (преимущественно путем миграции, выселения клеток из
У пресмыкающихся и их эволюционных потомков, птиц и млекопитающих, при переходе к размножению на суше наряду с зародышевыми оболочками
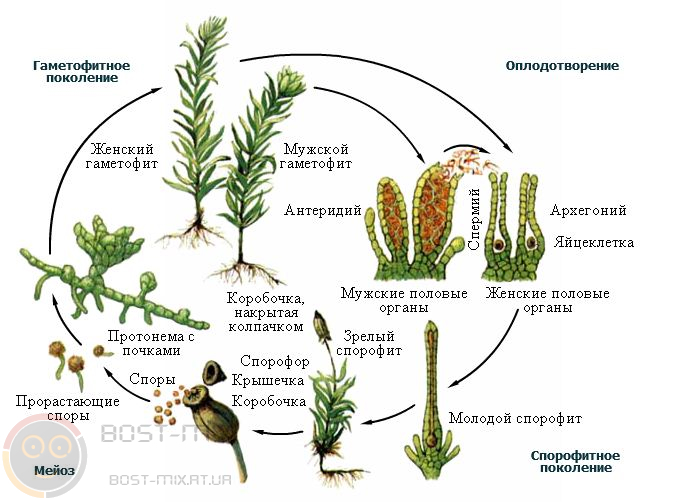
Постэмбриональное развитие бывает прямым и непрямым. При прямом развитии появившийся организм идентичен по строению взрослому организму, но имеет меньшие размеры и не обладает половой зрелостью. Дальнейшее развитие связано с увеличением размеров и приобретением половой зрелости. Такое развитие характерно для рептилий, птиц, млекопитающих. Непрямое развитие, или развитие с метаморфозом – появившийся организм отличается по строению от взрослого организма, может иметь специфические органы, и такой зародыш называется личинкой. Личинка питается, растёт, и со временем личиночные органы заменяются органами, свойственными взрослому организму (имаго). Развитие с личиночными стадиями характерно для бесхвостых амфибий (лягушки), насекомых, различных типов червей. Среди насекомых различают также насекомых с полным и неполным превращением (метаморфозом). Насекомые с неполным превращением проходят три стадии: яйцо, личинка, взрослая особь (имаго). Личинки насекомых с неполным превращением внешне сходны с имаго и, подобно последним, имеют сложные глаза, такой же, как у взрослых, ротовой аппарат и, в более старших возрастах, наружные зачатки крыльев. У многих насекомых с неполным превращением личинки ведут сходный с имаго образ жизни и могут встречаться совместно с последними. Развитие с неполным превращением характерно для стрекоз, тараканов, полужёсткокрылых (клопы), прямокрылых (кузнечики, саранча, цикады) и др. Полное превращение, характеризуется прохождением насекомым от четырех до пяти стадий – яйца, личинки, куколки, имаго, а иногда и предкуколки. Личинки насекомых с полным превращением, как правило, мало похожи на взрослых особей, поэтому называются истинными личинками. Действительно, трудно узнать в червеобразной личинке жука или в гусенице бабочку – взрослое крылатое насекомое. При этом личинка всегда лишена сложных глаз, наружных зачатков крыльев и часто имеет иной тип ротового аппарата, нежели взрослые особи. Обычно истинные личинки живут в иной, чем имаго, среде, и используют иные пищевые ресурсы. В связи с этим большинство органов истинных личинок имеют временный характер, после метаморфоза во время стадии куколки эти органы исчезают полностью и заменяются иными, характерными для имаго. Полное превращение характерно для отрядов двукрылых (комары, мухи), жёсткокрылых (жуки), чешуекрылых (бабочки), перепончатокрылых (пчелы, муравьи, осы). Причины нарушения развития организма могут иметь эндогенный – внутренний (мутации, хромосомные аномалии) и экзогенный – внешний – характер (действие радиации и некоторых химических веществ, нарушения обмена в материнском организме), причём, чем младше зародыш, тем чаще подобные нарушения оказываются несовместимыми с его жизнью. У растений жизненный цикл имеет более сложную форму, чем у животных, и сопряжён со сменою поколений полового (гаплоидного, гаметофита) и бесполого (диплоидного, спорофита). Эволюционной тенденцией в ряду
|